Información de la revista
- Enlace del artículo: 10.1002/jev2.70109
- Revista: Journal of Extracellular Vesicles
- Impact Factor: aproximadamente 25 (valor estimado)
- Acerca de la revista: La Journal of Extracellular Vesicles (JEV) es la revista oficial de la Sociedad Internacional de Vesículas Extracelulares (ISEV) y está especializada en la investigación sobre vesículas extracelulares (VE). Publica artículos de alta calidad que abarcan todos los aspectos de la investigación de las VE, incluidas la biología de las VE, la ingeniería de las VE y el diagnóstico y la terapéutica basados en VE, y se reconoce como una de las principales revistas en este campo.
Resumen
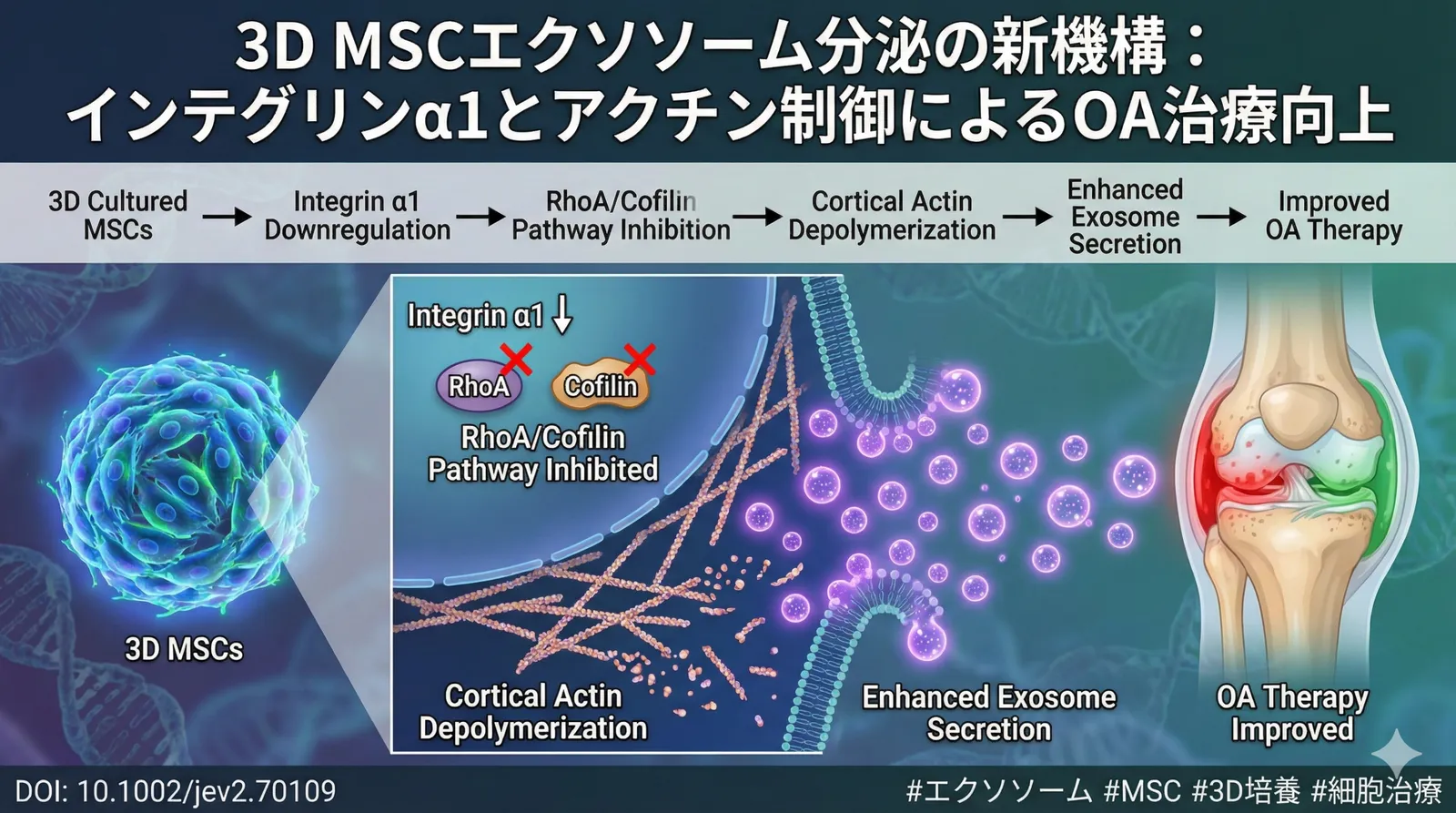
Este estudio tuvo como objetivo dilucidar, a nivel molecular, el mecanismo por el cual el cultivo tridimensional (3D) de células madre mesenquimales (CMM) promueve la secreción de vesículas extracelulares (sEV) en comparación con el cultivo bidimensional (2D). Como resultado, se reveló que en las CMM cultivadas en 3D, la regulación a la baja de la integrina α1 (ITGA1) suprime la vía de señalización RhoA/cofilina, lo que induce la despolimerización de la actina cortical y promueve la liberación de sEV. Además, se demostró que las sEV derivadas de CMM cultivadas en 3D potencian la eficacia terapéutica tanto in vitro como in vivo en modelos de rata de osteoartritis (OA) y de cicatrización de heridas. Este estudio identifica la despolimerización de la actina cortical dependiente de la vía RhoA/cofilina como un nuevo mecanismo que promueve la secreción de sEV, y aporta nuevas perspectivas para optimizar el rendimiento y la eficacia terapéutica de las sEV derivadas de células madre.
1. ¿Qué es exactamente la “actina cortical”?
Justo por dentro de la membrana celular, hay fibras proteicas llamadas “filamentos de actina” dispuestas en una red en forma de malla. A esto se le denomina **actina cortical (Cortical Actin)**.
- Función: Cumple la función de “armazón” que mantiene la forma de la célula y también la de “barrera física” que impide que las sustancias del interior de la célula salgan al exterior de manera incontrolada.
2. ¿Qué es la “despolimerización”?
A la disgregación de las fibras que están polimerizadas (convertidas en polímero) se le llama despolimerización. Es decir, es un estado en el que la malla de fibras de actina, que estaba firmemente ensamblada, se descompone y la malla se afloja hasta quedar dispersa.
3. ¿Por qué es esto importante para la “secreción de exosomas”?
Normalmente, los exosomas (y las vesículas que los contienen) del interior de la célula intentan salir a través de la membrana celular, pero la malla de actina cortical se interpone y les impide salir con facilidad.
Sin embargo, cuando se produce la **“despolimerización de la actina cortical”**:
- Desaparición de la barrera: La malla que obstruía se descompone y desaparece.
- Promoción de la fusión: Las vesículas que contienen exosomas pueden alcanzar la membrana celular sin obstáculos y fusionarse con ella.
- Aumento de la secreción: Como resultado, se liberan grandes cantidades de exosomas al exterior de la célula.
Antecedentes
Las vesículas extracelulares (VE) son mediadores importantes de la comunicación intercelular e influyen en procesos fisiológicos y patológicos al transportar sustancias bioactivas como proteínas, ácidos nucleicos y lípidos a las células diana. En particular, se ha informado que las vesículas extracelulares (sEV) derivadas de células madre mesenquimales (CMM) muestran diversos efectos terapéuticos, como reparación tisular, inmunomodulación y acción antiinflamatoria, y han atraído gran atención en el campo de la medicina regenerativa. Sin embargo, la cantidad de secreción de sEV por parte de las CMM es limitada, y se requiere el desarrollo de métodos eficaces para aumentar su rendimiento.
En comparación con el cultivo bidimensional (2D) convencional, los sistemas de cultivo tridimensional (3D) pueden aproximar la morfología celular, la expresión génica y las interacciones célula-célula a un estado más fisiológico, lo que se ha sugerido que conduce a una mejora de la función celular y a la promoción de la secreción de sEV. Sin embargo, el mecanismo molecular por el cual el cultivo 3D promueve la secreción de sEV por parte de las CMM no se había dilucidado completamente. Este estudio tiene como objetivo revelar un nuevo mecanismo en el que la despolimerización de la actina cortical promueve la secreción de sEV en las CMM cultivadas en 3D y, al analizar en detalle este mecanismo molecular, establecer una base para optimizar el rendimiento y la eficacia terapéutica de las sEV.
Laboratorio y autores
Este artículo fue publicado por el laboratorio del profesor Lim Chwee Teck de la Universidad Nacional de Singapur (National University of Singapore, NUS).
Acerca del laboratorio
El profesor Lim Chwee Teck está afiliado al Departamento de Ingeniería Biomédica y al Departamento de Ingeniería Mecánica de la Universidad Nacional de Singapur, y es también investigador principal del Mechanobiology Institute (MBI). El laboratorio Lim se centra en el papel de la señalización mecánica a nivel celular y tisular y, en particular, estudia la importancia de las fuerzas mecánicas en el cáncer, las enfermedades vasculares y la biología de las células madre. El laboratorio emplea una amplia variedad de tecnologías —incluidos los dispositivos de microfluídica, los biomateriales y las técnicas de imagen— para investigar cómo las células detectan estímulos mecánicos y responden a ellos. Sus hallazgos de investigación hasta la fecha se han publicado en revistas de primer nivel como Nature Materials, Nature Cell Biology y PNAS. Puede decirse que el presente artículo es una investigación en línea con uno de los temas de larga data del laboratorio: el efecto de los cambios del citoesqueleto en el cultivo celular 3D sobre la secreción de exosomas.
El laboratorio Lim se estableció en 1999 y desde entonces ha formado a numerosos investigadores. Según el sitio web del laboratorio (https://me.nus.edu.sg/bme/people/academic-staff/lim-chwee-teck/), los principales temas de investigación del laboratorio son los siguientes.
- Propiedades mecánicas de las células y el citoesqueleto: El laboratorio estudia cómo las propiedades mecánicas como la rigidez celular, la viscoelasticidad y la deformabilidad afectan a la función celular. En particular, se centra en los mecanismos por los cuales los componentes del citoesqueleto, como la actina, la miosina y los filamentos intermedios, controlan las propiedades mecánicas de las células.
- Interacciones entre la célula y la matriz extracelular: La matriz extracelular (MEC) funciona no solo como un andamiaje para las células, sino también como una importante molécula de señalización que controla la supervivencia, la proliferación y la diferenciación celulares. El laboratorio investiga cómo las células detectan las propiedades mecánicas y la composición de la MEC y responden a ellas. En particular, se centra en las interacciones entre la célula y la MEC mediadas por moléculas de adhesión celular como las integrinas.
- Manipulación celular mediante dispositivos de microfluídica: Dado que los dispositivos de microfluídica pueden controlar con precisión volúmenes minúsculos de líquido, constituyen una herramienta poderosa en la investigación de la biología celular. El laboratorio desarrolla tecnologías que utilizan dispositivos de microfluídica para medir las propiedades mecánicas de las células, aplicar estímulos mecánicos a las células y separar y concentrar células.
- Mecanobiología del cáncer: Se sabe que las células cancerosas exhiben propiedades mecánicas que difieren de las de las células normales. El laboratorio estudia cómo las propiedades mecánicas de las células cancerosas —como la rigidez, la deformabilidad y la capacidad invasiva— participan en la metástasis del cáncer y la resistencia a los fármacos. Asimismo, su objetivo es desarrollar nuevas terapias que tengan como diana las propiedades mecánicas de las células cancerosas mediante dispositivos de microfluídica.
- Mecanobiología de las células madre: Las células madre son células especiales con capacidad de autorrenovación y multipotencia, y se espera que tengan aplicaciones en la medicina regenerativa. El laboratorio estudia las propiedades mecánicas de las células madre y los mecanismos por los cuales las células madre se diferencian en respuesta a estímulos mecánicos. Asimismo, su objetivo es desarrollar tecnologías que utilicen dispositivos de microfluídica para controlar la diferenciación de las células madre.
Puede decirse que el presente artículo es una investigación en línea con uno de los temas de larga data del laboratorio: el efecto de los cambios del citoesqueleto en el cultivo celular 3D sobre la secreción de exosomas. En particular, se considera que se centra en cómo las interacciones con la matriz extracelular y las propiedades mecánicas de las células afectan a la secreción de exosomas.
Acerca de los autores
El autor de correspondencia (Corresponding Author) de este artículo es el profesor Lim Chwee Teck. Como se ha señalado anteriormente, el profesor Lim es un investigador destacado de la Universidad Nacional de Singapur y uno de los líderes en el campo de la bioingeniería. Su investigación combina la mecánica celular, la microfluídica y las aplicaciones biomédicas, y se centra en particular en el estudio del cáncer, las células madre y las vesículas extracelulares. El profesor Lim ha recibido numerosos premios por sus notables contribuciones al campo. Por ejemplo, recibió el Premio Presidencial de Ciencia y Tecnología de Singapur en 2016.
El profesor Lim ha publicado más de 400 artículos hasta la fecha, y sus logros gozan de un alto reconocimiento internacional. Su investigación ha tenido un gran impacto en los campos de la biología celular, los biomateriales y la medicina regenerativa, y se anticipa que continuará destacando en el futuro.
Antecedentes que condujeron a este estudio
El laboratorio Lim ha estudiado durante muchos años la relación entre las propiedades mecánicas de las células y la función celular. En particular, se ha centrado en cómo las interacciones con la matriz extracelular (MEC) y el estrés mecánico al que se someten las células afectan a la supervivencia, la proliferación, la diferenciación celulares y la secreción de vesículas extracelulares. Como parte de este trabajo, el presente estudio se centra en el efecto del cultivo celular 3D sobre las propiedades mecánicas de las células y analiza en detalle cómo este afecta a la secreción de exosomas. Considerando que el cultivo celular 3D está más cerca de un estado fisiológico que el cultivo celular 2D, el laboratorio ha hecho hincapié desde hace tiempo en la investigación de la función celular utilizando el cultivo celular 3D. El presente estudio es uno de los resultados de ese esfuerzo, y se considera que tiene un significado muy importante en el sentido de que dilucidó, a nivel molecular, el mecanismo por el cual el cultivo celular 3D promueve la secreción de exosomas.
Principales hallazgos – niveles molecular, celular y tisular
Resumen de los sistemas experimentales y los modelos animales
- Células utilizadas: Células madre mesenquimales derivadas de médula ósea humana (hMSC)
- Condiciones de cultivo: Cultivo 2D (matraz estándar de cultivo de tejidos) y cultivo 3D (andamio Alvetex®). Ambos cultivos utilizaron medio DMEM suplementado con un 10 % de FBS y un 1 % de penicilina/estreptomicina. Para el cultivo 3D, las células se sembraron en un andamio Alvetex® y se cultivaron en el mismo medio que el cultivo 2D.
- Modelo de osteoartritis (OA): Se indujo OA en ratas Sprague-Dawley macho de 6 semanas de edad mediante la inyección de monoyodoacetato (MIA) en la articulación de la rodilla derecha. La dosis de MIA fue de 2 mg/50 μL.
- Modelo de cicatrización de heridas: Se creó un defecto cutáneo de espesor completo de 8 mm de diámetro en el dorso de ratas Sprague-Dawley macho de 8 semanas de edad.
- Administración de sEV: En el modelo de OA y en el modelo de cicatrización de heridas, las sEV derivadas de hMSC cultivadas en 3D (100 μg/50 μL de PBS) se administraron localmente en el sitio de la lesión.
- Tamaño de la muestra: n = 5–8 por grupo experimental.
1. Explicación detallada a nivel molecular
1.1 Regulación a la baja de la integrina α1 (ITGA1)
En las hMSC cultivadas en 3D, la expresión de la integrina α1 (ITGA1) estaba significativamente reducida en comparación con el cultivo 2D, según se confirmó mediante Western blotting (Figure 1A). La ITGA1 es un receptor de la superficie celular que se une a componentes de la matriz extracelular (MEC) como el colágeno y la laminina, y desempeña un papel importante en funciones celulares como la adhesión celular, la migración celular y la diferenciación celular. En el entorno de cultivo 3D, se considera que las células modifican sus interacciones con la MEC y suprimen la expresión de ITGA1, induciendo así la reorganización del citoesqueleto. Concretamente, tras lisar las células, se extrajeron las proteínas, se separaron mediante electroforesis SDS-PAGE y se transfirieron a una membrana de PVDF. La detección se realizó utilizando un anticuerpo primario contra ITGA1 (p. ej., Abcam ab30393) y un anticuerpo secundario marcado con HRP. Las bandas se visualizaron mediante quimioluminiscencia y se analizaron cuantitativamente con el software ImageJ. Los niveles de expresión de ITGA1 se normalizaron con un gen constitutivo (p. ej., GAPDH).
1.2 Supresión de la vía de señalización RhoA/cofilina
La regulación a la baja de ITGA1 provoca la supresión de la vía de señalización RhoA/cofilina. RhoA es un tipo de GTPasa de bajo peso molecular implicada en la contracción del citoesqueleto de actina y en la regulación de la adhesión celular. La cofilina es una proteína que despolimeriza los filamentos de actina; es fosforilada por ROCK (Rho-associated kinase), que es activada por RhoA, y su actividad se regula de ese modo. En las hMSC cultivadas en 3D, los niveles de fosforilación de RhoA y de cofilina estaban significativamente reducidos, según se confirmó mediante Western blotting (Figure 1B, C). Esto sugiere que la regulación a la baja de ITGA1 suprime la activación de RhoA, lo que a su vez reduce la fosforilación de la cofilina y promueve la despolimerización de los filamentos de actina. La activación de RhoA puede evaluarse midiendo la cantidad de RhoA unida a GTP. La RhoA unida a GTP se separa del lisado celular mediante un ensayo de pull-down y se detecta por Western blotting. La fosforilación de la cofilina se detecta utilizando un anticuerpo específico de la cofilina fosforilada.
1.3 Promoción de la despolimerización de la actina
La supresión de la vía de señalización RhoA/cofilina promueve la despolimerización de la actina. La actina es un componente principal del citoesqueleto y participa en diversas funciones celulares como el mantenimiento de la morfología celular, la motilidad celular y el transporte intracelular. La actina adopta dos formas —la G-actina globular y la F-actina filamentosa— y permite la reorganización dinámica del citoesqueleto al someterse repetidamente a polimerización y despolimerización. En las hMSC cultivadas en 3D, la cantidad de F-actina disminuyó y la cantidad de G-actina aumentó, según se confirmó mediante microscopía de fluorescencia y Western blotting (Figure 2A, B). Esto sugiere que la supresión de la vía de señalización RhoA/cofilina promueve la despolimerización de los filamentos de actina, lo que hace que el citoesqueleto sea más flexible. La polimerización de la actina se evalúa utilizando faloidina marcada por fluorescencia. Dado que la faloidina se une específicamente a la F-actina, la observación por microscopía de fluorescencia permite evaluar la distribución y la cantidad de F-actina. También es posible lisar las células, separar la G-actina y la F-actina, y cuantificar la cantidad de cada una mediante Western blotting.
1.4 Supresión de la secreción de sEV mediante la inactivación (knockdown) de Rab27A/B
Rab27A y Rab27B son tipos de GTPasa de bajo peso molecular implicadas en el transporte de vesículas intracelulares, en particular en la secreción de vesículas extracelulares (VE). Se sabe que estas proteínas regulan el proceso por el cual las VE se fusionan con la membrana celular y se liberan al exterior de la célula. En este estudio, se crearon hMSC con Rab27A y Rab27B inactivadas (knockdown) y se examinó su efecto sobre la secreción de sEV. Los resultados revelaron que la inactivación de Rab27A y Rab27B suprimió significativamente la secreción de sEV (Figure 3A). Concretamente, la inactivación de Rab27A redujo la secreción de sEV a aproximadamente 0,5 veces, y la inactivación de Rab27B redujo la secreción de sEV a aproximadamente 0,1 veces. Este resultado sugiere que Rab27A y Rab27B desempeñan papeles esenciales en la secreción de sEV por parte de las hMSC. La inactivación de Rab27A/B se realiza utilizando siRNA o shRNA. Se introduce siRNA o shRNA en las hMSC para suprimir la expresión de Rab27A/B. La eficiencia de la inactivación se confirma mediante qRT-PCR o Western blotting.
1.5 Cambios en los niveles de expresión de Rab27A/B en el cultivo 2D/3D
Curiosamente, no se observó ninguna diferencia significativa en los niveles de expresión de Rab27A y Rab27B entre el cultivo 2D y el cultivo 3D (Figure 3B). Esto sugiere que la promoción de la secreción de sEV por el cultivo 3D no se debe a cambios en los niveles de expresión de Rab27A y Rab27B. Más bien, se considera que la despolimerización de la actina cortical inducida por el cultivo 3D promueve la secreción de sEV a través de una vía independiente de Rab27A/B. En la qRT-PCR se mide la cantidad de ARNm de Rab27A/B. Se extrae el ARN total, se retrotranscribe a ADNc y se realiza la PCR utilizando cebadores específicos de Rab27A/B. Los niveles de expresión se normalizan con un gen constitutivo (p. ej., GAPDH). En el Western blotting, la cantidad de expresión proteica se mide utilizando anticuerpos contra Rab27A/B.
2. Explicación detallada a nivel celular
2.1 Cambios morfológicos en las hMSC
Se confirmó mediante microscopía de contraste de fase que las hMSC cultivadas en 3D presentan una morfología más cercana a la esférica en comparación con el cultivo 2D (Figure 4A). Esto sugiere que, en el entorno de cultivo 3D, las células debilitan su adhesión al andamio y reorganizan su citoesqueleto, adoptando así una morfología más libre. La morfología celular se observa utilizando microscopía de contraste de fase o microscopía láser confocal. Tras fijar las células, se tiñen la membrana celular y el citoesqueleto y se observan al microscopio. En el cultivo 3D también es posible observar cómo las células se infiltran en el interior del andamio.
2.2 Reorganización del citoesqueleto de actina
En las hMSC cultivadas en 3D, se confirmó mediante microscopía de fluorescencia que el citoesqueleto de actina se reorganizaba de forma más flexible (Figure 4B). Mientras que en el cultivo 2D los filamentos de actina se disponen en paralelo a lo largo de la membrana celular, en el cultivo 3D se observó que los filamentos de actina se disponían de forma más aleatoria y se distribuían de manera uniforme por toda la célula. Esto sugiere que la despolimerización de la actina cortical inducida por el cultivo 3D aumenta la flexibilidad del citoesqueleto y promueve los cambios en la morfología celular y la motilidad celular. La visualización del citoesqueleto de actina se realiza utilizando faloidina marcada por fluorescencia. Dado que la faloidina se une específicamente a la F-actina, la observación por microscopía de fluorescencia permite observar en detalle la distribución y la estructura de los filamentos de actina.
2.3 Aumento de la cantidad de sEV liberada
Se confirmó mediante análisis de seguimiento de nanopartículas (NTA) que las hMSC cultivadas en 3D liberan una cantidad significativamente mayor de sEV en comparación con el cultivo 2D (Figure 5A). El NTA es una técnica para medir el tamaño y la concentración de micropartículas (como las sEV) en un líquido y se utiliza ampliamente en la investigación de las sEV. El hecho de que el cultivo 3D aumentara la cantidad de sEV liberada en aproximadamente 2 veces sugiere que el cultivo 3D es un medio eficaz para mejorar la eficiencia de producción de sEV. La cantidad de sEV liberada se mide recogiendo el sobrenadante del cultivo, eliminando las células y los restos (debris) por centrifugación, y midiéndola luego mediante NTA. En el NTA se aplica luz láser y se analiza la luz dispersada por las micropartículas para calcular el tamaño y la concentración de las partículas.
2.4 Cambios en la captación de sEV
Las sEV son captadas por las células diana a través de mecanismos como la endocitosis, y median la comunicación intercelular al transportar sustancias bioactivas —como las proteínas y los ácidos nucleicos contenidos en su interior— a las células diana. Se confirmó mediante microscopía de fluorescencia y citometría de flujo que las sEV derivadas de hMSC cultivadas en 3D se captaban en mayores cantidades por las células diana (por ejemplo, condrocitos y fibroblastos) en comparación con las sEV derivadas de hMSC cultivadas en 2D (Figure 5B). Este resultado sugiere que el cultivo 3D potencia la capacidad de captación celular de las sEV, lo que podría contribuir a mejorar la eficacia terapéutica de las sEV. La capacidad de captación de las sEV se evalúa utilizando sEV marcadas por fluorescencia. Las sEV se marcan con un colorante fluorescente (por ejemplo, DiI o CFSE), se añaden a las células diana y se cultivan durante un tiempo determinado. A continuación, las células se lavan y se mide la cantidad de sEV captadas en el interior de las células mediante microscopía de fluorescencia o citometría de flujo.
3. Explicación detallada a nivel tisular
3.1 Efecto protector del cartílago en el modelo de osteoartritis (OA)
La osteoartritis (OA) es una enfermedad articular crónica caracterizada por la degeneración y la destrucción del cartílago articular, y es uno de los factores que reducen notablemente la calidad de vida (QOL) de las personas mayores. En este estudio, cuando se administraron sEV derivadas de hMSC cultivadas en 3D a un modelo de rata con OA, se suprimió la destrucción del cartílago y mejoró la tasa de supervivencia de los condrocitos, según se confirmó mediante análisis histológico (tinción con hematoxilina y eosina, tinción con safranina O y tinción con azul de toluidina) (Figure 6A, B). Estos resultados sugieren que las sEV derivadas de hMSC cultivadas en 3D tienen un efecto de supresión de la progresión de la OA y de protección del cartílago. En el análisis histológico, se extrae el tejido articular, se incluye en parafina, se corta en secciones finas y se somete a diversas tinciones. La tinción con hematoxilina y eosina permite observar la morfología celular y la estructura tisular. La tinción con safranina O y la tinción con azul de toluidina permiten evaluar la cantidad de proteoglicano del cartílago. El grado de destrucción del cartílago se cuantifica utilizando criterios de evaluación como la puntuación OARSI.
3.2 Efecto promotor del cierre de la herida en el modelo de cicatrización de heridas
La cicatrización de heridas es un proceso complejo que repara el daño cutáneo y progresa a través de etapas como la inflamación, la proliferación celular y la remodelación tisular. En este estudio, cuando se administraron sEV derivadas de hMSC cultivadas en 3D a un modelo de rata de cicatrización de heridas, se promovió el cierre de la herida y se promovió la formación de tejido de granulación, según se confirmó mediante observación macroscópica y análisis histológico (tinción con hematoxilina y eosina y tinción tricrómica de Masson) (Figure 7A, B). Estos resultados sugieren que las sEV derivadas de hMSC cultivadas en 3D tienen un efecto de promoción de la cicatrización de heridas. El grado de cierre de la herida se evalúa midiendo el área de la herida. La formación de tejido de granulación se observa mediante tinción con hematoxilina y eosina, y se evalúan el grado de angiogénesis y la cantidad de depósito de colágeno. La tinción tricrómica de Masson permite observar en detalle la distribución del colágeno.
4. Explicación detallada de los resultados de verificación en modelos animales
4.1 Modelo de osteoartritis (OA)
- Modelo animal: Ratas Sprague-Dawley macho de 6 semanas de edad
- Inducción de OA: Inyección de monoyodoacetato (MIA) en la articulación de la rodilla derecha (2 mg/50 μL)
- Administración de sEV: sEV derivadas de hMSC cultivadas en 3D (100 μg/50 μL de PBS) administradas en la cavidad de la articulación de la rodilla dos veces por semana, cuatro veces en total
- Evaluación: Análisis histológico (tinción con hematoxilina y eosina, tinción con safranina O, tinción con azul de toluidina) realizado a las 4 semanas tras la administración
- Resultados: En el grupo al que se administró sEV, la destrucción del cartílago debida a la administración de MIA se suprimió significativamente y mejoró la tasa de supervivencia de los condrocitos
- Análisis estadístico: ANOVA followed by Tukey’s post-hoc test
4.2 Modelo de cicatrización de heridas
- Modelo animal: Ratas Sprague-Dawley macho de 8 semanas de edad
- Creación de la herida: Se creó un defecto cutáneo de espesor completo de 8 mm de diámetro en el dorso
- Administración de sEV: sEV derivadas de hMSC cultivadas en 3D (100 μg/50 μL de PBS) administradas localmente en el sitio de la herida
- Evaluación: Se midió la tasa de cierre de la herida a los 14 días tras la administración y se realizó un análisis histológico (tinción con hematoxilina y eosina, tinción tricrómica de Masson)
- Resultados: En el grupo al que se administró sEV, el cierre de la herida se promovió significativamente y se promovió la formación de tejido de granulación
- Análisis estadístico: ANOVA followed by Tukey’s post-hoc test
5. Interpretación específica de los datos experimentales
5.1 Figure 1: Análisis de la vía ITGA1 y RhoA/cofilina
- Figure 1A: Niveles de expresión de ITGA1 en hMSC cultivadas en 2D y en 3D comparados mediante Western blotting. En el cultivo 3D, la expresión de ITGA1 estaba significativamente reducida (p < 0.05, Student’s t-test).
- Figure 1B: Niveles de expresión de p-RhoA (RhoA fosforilada) en hMSC cultivadas en 2D y en 3D comparados mediante Western blotting. En el cultivo 3D, la expresión de p-RhoA estaba significativamente reducida (p < 0.05, Student’s t-test).
- Figure 1C: Niveles de expresión de p-cofilina (cofilina fosforilada) en hMSC cultivadas en 2D y en 3D comparados mediante Western blotting. En el cultivo 3D, la expresión de p-cofilina estaba significativamente reducida (p < 0.05, Student’s t-test).
5.2 Figure 2: Análisis de la despolimerización de la actina
- Figure 2A: Distribución de la F-actina en hMSC cultivadas en 2D y en 3D observada mediante microscopía de fluorescencia. En el cultivo 3D, la F-actina disminuyó y se distribuyó de manera uniforme por toda la célula.
- Figure 2B: Relación F/G-actina en hMSC cultivadas en 2D y en 3D cuantificada mediante Western blotting. En el cultivo 3D, la relación F/G-actina estaba significativamente reducida (p < 0.05, Student’s t-test).
5.3 Figure 3: Efecto de la inactivación (knockdown) de Rab27A/B
- Figure 3A: Cantidad de secreción de sEV en hMSC con Rab27A/B inactivadas medida mediante análisis de seguimiento de nanopartículas (NTA). La inactivación de Rab27A y Rab27B redujo significativamente la secreción de sEV (p < 0.01, ANOVA followed by Tukey’s post-hoc test).
- Figure 3B: Niveles de expresión de Rab27A/B en hMSC cultivadas en 2D y en 3D medidos mediante qRT-PCR. Sin diferencia significativa en los niveles de expresión de Rab27A/B entre el cultivo 2D/3D.
5.4 Figure 6: Efecto protector del cartílago en el modelo de OA
- Figure 6A: Tejido de la articulación de la rodilla del modelo de rata con OA observado mediante tinción con hematoxilina y eosina. En el grupo al que se administró sEV, la destrucción del cartílago se suprimió.
- Figure 6B: Tejido de la articulación de la rodilla del modelo de rata con OA observado mediante tinción con safranina O. En el grupo al que se administró sEV, mejoró la tinción del proteoglicano.
5.5 Figure 7: Efecto promotor del cierre de la herida en el modelo de cicatrización de heridas
- Figure 7A: Sitio de la herida del modelo de rata de cicatrización de heridas observado macroscópicamente. En el grupo al que se administró sEV, el cierre de la herida se promovió.
- Figure 7B: Sitio de la herida del modelo de rata de cicatrización de heridas observado mediante tinción con hematoxilina y eosina. En el grupo al que se administró sEV, la formación de tejido de granulación se promovió.
Discusión / Implicaciones
Perspectiva del antienvejecimiento
Se sabe que las células madre mesenquimales (CMM) declinan en su función con el envejecimiento. El mecanismo de promoción de la secreción de sEV por el cultivo 3D demostrado en este estudio puede convertirse en una nueva estrategia para mejorar la capacidad de secreción de sEV de las CMM envejecidas y potenciar los efectos antienvejecimiento. Por ejemplo, se espera que su combinación con la tecnología de cultivo 3D potencie la eficacia terapéutica de las sEV derivadas de CMM envejecidas. El control dinámico del citoesqueleto de actina es importante para mantener la juventud de las células, y los fármacos o los compuestos de moléculas pequeñas que tengan como diana la vía RhoA/cofilina pueden convertirse en candidatos para la terapia antienvejecimiento.
Perspectiva de la medicina regenerativa (CMM / EV)
Las sEV derivadas de CMM están atrayendo atención en el campo de la medicina regenerativa como una nueva terapia que puede sustituir al trasplante celular. Este estudio mostró que el cultivo 3D mejora la eficiencia de producción de sEV y potencia la eficacia terapéutica. Esto puede considerarse un logro importante para acelerar la aplicación práctica de la medicina regenerativa que utiliza sEV. En particular, se anticipa el desarrollo de la terapia con sEV para enfermedades crónicas como la osteoartritis y la cicatrización de heridas. Además, puede esperarse una mejora adicional de la eficacia terapéutica optimizando el método de administración, la dosis y el intervalo de dosificación de las sEV.
Perspectiva de la interacción nervio–órgano
Se ha sugerido que las sEV pueden mediar la comunicación entre el sistema nervioso y otros órganos. La eficacia terapéutica de las sEV demostrada en este estudio puede reflejar un mecanismo de reparación tisular mediado por la interacción nervio–órgano. Por ejemplo, las sEV podrían actuar sobre las células del sistema nervioso para promover la secreción de factores neurotróficos o suprimir la inflamación, promoviendo así indirectamente la reparación tisular. En investigaciones futuras será importante analizar en detalle los efectos de las sEV sobre el sistema nervioso y dilucidar la eficacia terapéutica de las sEV mediada por la interacción nervio–órgano.
Perspectivas de futuro
Este estudio dilucidó el mecanismo por el cual el cultivo 3D promueve la secreción de sEV derivadas de CMM y abrió el camino al desarrollo de nuevas terapias que utilizan sEV. En investigaciones futuras serán importantes los siguientes puntos.
- Aplicación clínica: Sobre la base de los hallazgos obtenidos en este estudio, será necesario realizar ensayos clínicos de la terapia con sEV para enfermedades crónicas como la osteoartritis y la cicatrización de heridas, y evaluar su seguridad y eficacia. Además, puede esperarse una mejora adicional de la eficacia terapéutica optimizando el método de administración, la dosis y el intervalo de dosificación de las sEV.
- Control de calidad de las sEV: Dado que la calidad de las sEV afecta en gran medida a la eficacia terapéutica, será necesario establecer tecnologías de control de calidad para las sEV. Concretamente, el tamaño, la concentración, la composición proteica y la composición de ácidos nucleicos de las sEV deben controlarse estrictamente para minimizar la variación entre lotes.
- Direccionamiento de las sEV: La administración selectiva de las sEV a las células diana es un desafío importante para potenciar la eficacia terapéutica. Al modificar la superficie de las sEV con moléculas específicas, se espera poder potenciar la capacidad de unión de las sEV a las células diana y mejorar la eficacia terapéutica.
- Medicina personalizada: Se desea la realización de una medicina personalizada en la que se seleccione la terapia con sEV óptima de acuerdo con el estado de la enfermedad y los antecedentes genéticos del paciente. Concretamente, al analizar las muestras de sangre y de tejido del paciente y predecir la sensibilidad a las sEV, será posible proporcionar un tratamiento más eficaz.
Conclusión
Este estudio reveló que en las CMM cultivadas en 3D, la regulación a la baja de ITGA1 suprime la vía de señalización RhoA/cofilina, lo que induce la despolimerización de la actina cortical y promueve la liberación de sEV. Además, se demostró que las sEV derivadas de CMM cultivadas en 3D potencian la eficacia terapéutica en modelos de rata de OA y de cicatrización de heridas. Estos resultados identifican la despolimerización de la actina cortical dependiente de la vía RhoA/cofilina como un nuevo mecanismo que promueve la secreción de sEV, y aportan nuevas perspectivas para optimizar el rendimiento y la eficacia terapéutica de las sEV derivadas de células madre. Se anticipa que las investigaciones futuras acelerarán la aplicación práctica de la medicina regenerativa que utiliza sEV y proporcionarán a muchos pacientes nuevas opciones de tratamiento.